The present project represents an innovative means to assess student mastery of course materials within a relatively short time period. High school students were required to learn names for different countries as part of a geography assignment. A technique that combines electrical engineering and event related potentials (ERPs) monitored their learning and indicated after a 15 minute period how successful these students were in learning to associate names with the outlines of different countries. Comparisons of the ERP brain wave data with students' actual test scores showed high agreement between the two measures. In addition, the brain responses discriminated between when the material was mastered versus when students were only familiar with the material but had not yet mastered it.
The present paper outlines an attempt to use a neuroelectrical technique, event related potentials (ERPs) to study learning in young adolescents while they are engaged in a simulated geography lesson. This approach assumes that a relatively direct relationship exists between cognitive processes and changes in the brain's electrical activity. The ERP is a portion of the ongoing EEG activity of the brain that is specifically time-locked to some external or internal event. ERPs are complex waveforms thought to reflect changes in brain activity via fluctuations in various portions of this waveform. The wave is made up of a series of positive and negative peaks that will vary in peak latency (delay from stimulus onset) or amplitude (height of the wave) at different points during the time course of the wave following some stimulus event (Callaway, et al., 1978). Because of this time-locked feature, ERPs can reflect both general and specific aspects of the evoking stimulus (Molfese, 1978a, 1978b) as well as the individual's perceptions and decisions regarding the stimulus (Molfese, 1983; Nelson & Salapatek, 1986; Ruchkin, et al., 1981).
A number of studies have previously utilized this technology to study either directed learning (Holamon, Morris, & Retzlaff, 1995, Honda, Barrett, Yoshimura, Ikeda, Nagamine, & Shibasaki, 1996; Hugdahl & Nordby, 1991; van Hooff, Brunia, & Allen, 1996) or incidental learning (Noldy, Stelmack, & Carnpbell, 1990; Paller, Kutas, & Mayes, 1987; Paller, McCarthy, & Wood, 1988). Much of this work has focused on two regions of the ERP that appear to vary with learning - the P300 component and the late portion of the ERP between 400 and 800 ms following stimulus onset. The P300 usually refers to the third large positive peak in the ERP that occurs with a peak latency of approximately 300 ms after stimulus onset.
Holamon, Morris, & Retzlaff(1995) examined P300 amplitude and latency during a recognition task. Twenty-four adults first learned pairs of words from the WMS-R Verbal Paired Associate Learning Task. In a subsequent test, some of these pairs were again presented while others were broken up and recombined with other words. Thus, adults sometimes saw the original training pairs while on other trials they viewed mismatched pairs. Holamon et al. found that the P300 peak amplitude was greater during adult recognition of matching word pairs. When words within a pair did not match the original training set, the P300 was smaller in amplitude. This effect was noted to be most prominent at central and parietal recording sites (i.e., at the top and towards the back sides of the head) while the P300 peak latency (i.e., the point at which the peak was most positive) was shorter for matching than for non-matching pairs. Thus, under matching conditions when adults noted that the two words had been previously learned together, the P300 actually occurred earlier in time, perhaps suggesting more ready processing of the material due to previous learning.
A similar finding was reported by van Hooff, Brunia, & Allen ( 1996). In three recognition tasks, administered on two consecutive days, subjects were instructed to indicate when they saw recently learned words. These words were presented along with unlearned words and along with previously learned words which both required a non-recognition response. Recently learned target words as well as previously learned non-target words elicited a centro-parietal positivity between 500 and 1000 ms post-stimulus. As in the case of Holamon, Morris, & Retzlaff (1995), the results suggest that ERPs are sensitive to differences between learned and unlearned words.
As noted briefly above, material mastered during incidental learning tasks also produce changes in the ERP. Paller, McCarthy, & Wood (1988) subjects viewed words under incidental learning conditions in which each word required a two-choice decision based on semantic criteria (interesting/uninteresting or edible/inedible). Memory for those words was subsequently assessed in a free recall test followed by a recognition test. Event-related brain potentials elicited in response to the original presentation of each word were found to differ as a function of later memory performance. ERP responses to remembered words were more positive between 400 and 800 ms relative to the ERPs elicited in response to forgotten words. An earlier incidental study by Paller, Kutas, & Mayes ( 1987) also reported a late positive ERP that was elicited by words later recalled or recognized. This response was larger than that elicited by words that were apparently later forgotten and not recalled or recognized.
Subsequent studies have investigated incidental learning not only for words but for pictures as well. Noldy, Stelmack, & Campbell (1990) recorded ERPs during intentional or incidental learning of pictures and words, as well as during the subsequent recognition memory test for these stimuli. As expected, intentionally learned pictures were remembered better than incidentally learned pictures and intentionally learned words, which, in turn, were remembered better than incidentally learned words. The ERP results appeared to mirror these behavioral results. The pictures that were attended produced greater positive ERP amplitudes frontally at 250 ms and centro-parietally at 350 ms. Greater negativity at 450 ms at parietal and occipital sites was also noted in response to attended over non-attended pictures.
Honda, Barrett, Yoshimura, Ikeda, Nagamine, & Shibasaki, (1996) studied event-related potentials (ERPs) recorded from 21 scalp electrodes during a visual paired association task with delayed discrimination in 10 normal right-handed subjects. After adults completely memorized four pairs of figures during a learning period, they were presented with each cue (S 1) and asked to judge whether the following figure (S2) formed one of the memorized pairs or not. A choice reaction task with delay was used for control in an attempt to identify brain activity specifically related to memory function. Two ERP components associated with memory function were identified in the difference waveform between the responses in the paired association task and those in the choice reaction task. One ERP component was characterized by marked positivity over posterior recording sites between 390 ms and 1100 ms after the S 1 presentation. The second ERP component was a sustained frontal negative component which began at about 1100 ms after S I and ended just before the S2 presentation.
Both potentials were distributed predominantly on the scalp overlying the left hemisphere. Honda et al interpreted these findings to indicate that the early posterior positive component reflected retrieval of information from association memory, whereas the late frontal negativity reflected retention of information in working memory. Somewhat similar left hemisphere effects in a learning situation have also been noted by Hugdahl & Nordby (1991).
Procedure: This study consisted of three parts: (1) a pretraining session during which 6 male and 6 female right handed 16 - 17 years old adolescents learned new names for a set of new countries as part of a geography lesson. During this pretraining part, students viewed a series of novel names and outlines of "new" countries. On each trial a nonsense word appeared on a computer screen for 2 seconds, followed briefly by a blank screen, and then by a 2 second display of a randomly generated shape. Students pressed one of two computer keys to indicate whether a name which appeared with a picture was the actual name for that picture (a match) or not (a mismatch). Since no training occurred during this session, behavioral responses were expected to be at chance levels. (2) The second part involved a 15 minute training period during which students were trained to a no-error criterion for half of the names and shapes while the other half of the names and shapes were also presented for the same time period but were not paired together and not trained. (3) The third phase was a post-training period where names and pictures were presented in exactly the same fashion as in the pretraining trials. The stimuli consisted of twenty different shapes controlled for area and 20 different CVCVC nonsense names controlled for frequency and order of letter occurrence. All stimulus pairing and response keys were counterbalanced across conditions and students. Visual ERPs (VERPs) were recorded to the nonsense shapes during both the pre- and post-training sessions from scalp electrodes placed over left and right frontal (F3, F4), central-temporal (T3, T4), and parietal (P3, P4) scalp regions. These electrodes were referred to linked ear references (A 1, A2).
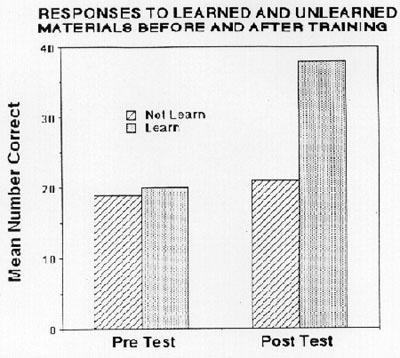
Results: Analyses of variance with Greenhouse-Geisser corrections evaluated the behavioral and VERP data. A significant PRE/POST x LEARNED/NOT-LEARNED interaction for the behavioral responses, F(1,11) = 33.76, p<.0001, indicated that performance improved for only the trained stimuli from pre- to post-training, F(l, l 1) = 99.54, p<.0001. As indicated in the figure below, there were no differences in the number of correct responses made during the pretest situation before training had occurred. For both lists of materials, the students responded at chance levels, only making on average about 20 correct responses out of 40 (chance = 50%). Performance during the post test showed a marked increase for the material learned during the preceding 15 minute training period. However, behavioral responses continued at chance levels for the material that had not been trained
Following standard artifact rejection and analysis procedures, the VERPs were averaged separately for the pre and post training conditions (2), for the learned and not learned stimulus sets (2), for the match and mismatch conditions (2), for the three electrode sites (3) over each hemisphere (2). Area under the curve measures were calculated using the EPACS (Molfese 1989) software package. Area under the curve measures of the P300 and late positive component (LPC) components thought to reflect cognitive processing of the 576 averaged visual VERPs between 350 ms and 850 ms following stimulus onset for area yielded a PRE/POST x LEARNED/NOT-LEARNED interaction, F(2,22) = 5.25, p<.043, indicating that ERPs differed as a function of the training. Area measures increased significantly for the P300 when VERPs were recorded to the learned materials during the post-training situation over the response to these same materials during the pretraining session, F(1,22) = 8.76, p<.013. No difference in area was noted for the VERPs elicited by the untrained materials. In addition, a PRE/POST x LEARNED/NOT-LEARNED x ELECTRODE interaction, F(2,22) = 3.479, p<.048, indicated that VERPs in the P300 region recorded over frontal and parietal sites during the post-training condition discriminated learned from not learned stimuli. The LPC region of the VERPs, however, changed for both the trained and untrained stimuli from the pretraining to the post-training trials, suggesting that this late occurring ERP component reflected a familiarization effect. These effects are illustrated in the second figure below.
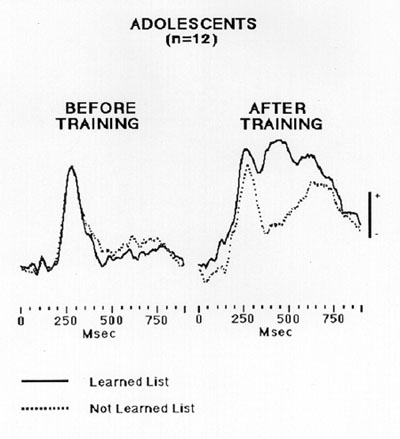
In this case, the two group averaged VERPs elicited in response to the country outlines prior to training are displayed on the left side of the figure. VERPs elicited to the two stimulus sets are virtually identical. Both waveforms are characterized by a single large positive peak that begins to increase in amplitude approximately 20 ms following stimulus onset, peaks at approximately 300 ms, and declines to minimal levels by 450 ms. The two VERPs on the right side of the figure, however, differ markedly from each other. While both waveforms increase at approximately the same time (175 ms) and reach their most positive early peak at approximately 260 ms, the VERP elicited in response to the untrained material (dashed line) shows a marked negative going wave that peaks at approximately 375 ms, that is then followed by a slow late positive component (LPC) that peaks at approximately 650 ms and then begins to decline again. The group averaged VERP elicited in response to the trained materials (solid line) shows a marked positive peak at approximately 400 ms post stimulus onset and then later a second late positive peak at 625 ms before declining in amplitude to parallel and overlap the VERP elicited by the untrained material. This distinction between the VERPs elicited during the post test session by the trained and untrained material reflects, we believe, a difference in the VERPs to familiar versus learned material. Both VERPs during the post test show a LPC. This could signal familiarity. However, the VERP elicited by the learned material is uniquely characterized by the large positive peak that begins at approximately 300 ms and peaks at approximately 400 ms post stimulus onset. This P400 behaves similarly to the P300-P400 effects reported by other investigators using both directed (Holamon, Morris, & Retzlaff, 1995; Honda, Barrett, Yoshimura, Ikeda, Nagamine, & Shibasaki, 1996; Hugdahl & Nordby, 1991; van Hooff, Brunia, & Allen, 1996) and incidental learning tasks (Noldy, Stelmack, & Campbell, 1990; Paller, Kutas, & Mayes, 1987; Paller, McCarthy, & Wood, 1988).
ERPs appear to be a reliable tool that can detect differences in learning after a relatively short time. In the present study, marked differences were noted in the VERP waveforms after only 15 minutes. Moreover, the technique may be able to differentiate effectively familiarity versus a more extensive mastery of the material. While the behavioral test (as illustrated in the first figure) could only indicate a performance increase for the learned material from the pre test to the post training test, the VERPs could apparently show that the students were in fact able to process the unlearned material differently following the training period. It would appear that the brain response could indicate the level at which the student has processed the material. This technique, then, may offer a sensitive and effective means to monitor the student's mastery of course material. Future research, however, is needed to determine whether this technique may differentiate degrees of mastery as well as the extent that it can differentiate familiar from novel from learned material under different types of learning conditions and with different types of materials .
Summary: ERPs reliably discriminated familiar from learned material as evidenced by marked increases in the area of the late occurring ERP regions, P300 and the LPC, for the learned versus the unlearned material during the post-training period. This technique may be useful for monitoring learning and perhaps even the identification of teaching techniques and curriculum material which are better suited to students of different levels of skill as abilities.
Callaway, C., Tueting, P., & Koslow, S. ( 1978). Event-related brain potentials and behavior. New York: Academic Press.
Holamon, B., Morris, G., & RetzlaffP. (1995). Event-related potentials during delayed recognition of Wechsler Memory Scale-R paired associate learning. Journal of Clinical Psychology. 51(3),39 1-5.
Hugdahl, K., & Nordby, H. (1991). Hemisphere differences in conditional learning: an ERP-study. Cortex. 27(4),557-70.
Honda, M., Barrett, G., Yoshimura, N., Ikeda, A., Nagamine, T., & Shibasaki, H. (1996). Event-related potentials during paired associate memory paradigm. Electroencephalography & Clinical Neurophysiology. 100(5),407-21
Molfese, D. (1978a). Electrophysiological correlates of categorical speech perception in adults. Brain and Language. 5, 25-35.
Molfese, D. (1983). Event related potentials and language processes. In A. Gaillard and W. Ritter (Eds.). Tutorials in ERP Research - Endogenous Components. Holland: Elsevier.
Molfese, D. (1978b). Left and right hemisphere involvement in speech perception: Electrophysiological correlates. Perception and Psychophysics. 23, 237-243.
Nelson, C., & Salapatek, P. (1986). Electrophysiological correlates of infant recognition memory. Child Development. 57, 1483- 1497.
Noldy, N.E., Stelmack, R.M., & Campbell, K.B. (1990). Event-related potentials and recognition memory for pictures and words: the effects of intentional and incidental learning. Psychophysiology. 2 7(4),417-28.
Paller, K.A., McCarthy, G., & Wood, C.C. (1988). ERPs predictive of subsequent recall and recognition performance. Biological Psychology. 26(1-3),269-76.
Paller, K.A., Kutas, M. & Mayes, A.R. (1987). Neural correlates of encoding in an incidental learning paradigm. Electroencephalography & Clinical Neurophysiology. 67(4),360-71.
Ruchkin, D., Sutton, S., Munson, R., & Macar, F. (1981). P300 and feedback provided by the absence of the stimuli. Psychophysiology. 18, 271-282.
van Hooff, J.C., Brunia, C.H., & Allen, J.J. (1996). Event-related potentials as indirect measures of recognition memory. International Journal of Psychophysiology. 21 (1), 15-31
 Back to Table of
Contents
Back to Table of
Contents